Mikoryza (od greckiego MójKomp „grzyba” i rhiza , „root”, to termin wprowadzony w 1885 roku przez botanika Albert Bernhard Frank ) jest wynikiem symbiotycznej stowarzyszenia o nazwie mikoryzacja , pomiędzy grzybów i korzeni z roślin . Mikoryza jest głównym składnikiem edafonu i ryzosfery .
W tym zazwyczaj niespecyficzne związku, gdy zarodniki o Mikoryzowego lub mikoryzowymi grzyba (od greckiego MójKomp , rhiza i genos „aby wywołać”, dosłownie, które przyczyniają się do mikoryzie) są rozprowadzane przez wiatr ( anemochoria ) przez deszcz ( hydrochorie ) lub odchodów zwierzęcych ( endozoochory ), kiełkowania i dawania dojścia do w grzybni z grzybni , która kolonizuje korzenie roślin. Te strzępki korzeni różnią się od strzępek pozakorzeniowych, które również rozwijają się poza korzeniem przez kilka centymetrów, badając glebę wokół systemu korzeniowego rośliny żywicielskiej. To, co powszechnie nazywa się grzybem, który zbiera się łapą i kapeluszem, jest jedynie „owocowaniem” grzybni, sporoforem , w którym następuje rozmnażanie płciowe . Strzępki wyglądają jak cienkie włókna, zdolne do eksploracji bardzo dużej objętości gleby (tysiąc metrów włókien grzybni na jeden metr korzenia).
Związek mikoryzowy jest typu symbiotycznego , ale brak równowagi w związku może być wywołany słabością jednego z dwóch partnerów, a związek może wtedy przesuwać się wzdłuż kontinuum mutualizmu i pasożytnictwa. Grzyb może wtedy również pomóc w recyklingu nekromasy swojego żywiciela, z korzyścią dla ich dwóch potomków.
O ile powierzchnia eksploracyjna wykorzystywana przez roślinę jest zwielokrotniana dziesięciokrotnie przez aktywne włośniki tylko w okresie kiełkowania, o tyle jest zwielokrotniana przez 10000 dzięki mikoryzom (filamenty o średnicy jednej setnej milimetra), które mają koszt energetyczny zawiązania sto razy mniej niż korzenie. Włośniki pozostają funkcjonalne w kilku grupach dorosłych roślin nie mikoryzowych (mniej niż 10%), które wtórnie utraciły asocjację grzybową, ponad 90% jest skolonizowanych przez te strzępki symbionta, które zapewniają większość wchłaniania wody i składników odżywczych oraz dziesięciokrotnie zwiększają wzrost roślin. Szacunki sugerują, że istnieje blisko 50 000 gatunków grzybów, które tworzą powiązania mikoryzowe z 250 000 gatunkami roślin, przy czym 80% tych powiązań to endomikoryzy arbuskularne. Wydaje się, że rośliny niemikoryzowe ze środowisk pionierskich (brak grzybów), wilgotne i/lub bogate, w których zaopatrzenie hydromineralne nie wymaga mikoryz, „musiały zrekompensować brak grzybiczej substancji pomocniczej poprzez rozwój struktur korzeniowych naśladujących grzybnię. włókna i ich wysoka skuteczność w kolonizowaniu dużej objętości gleby ”.
Aktualne badania wskazują na szerszy związek na poziomie mikoryz z pojęciem mikrobiomu mikoryzowego, zwanego rizomikrobiomem, który obejmuje również bakterie.
Oryginalny symbiont (a glomeromycete to wydaje), że pojawiły się około 500 milionów lat temu w paleozoiku , prawdopodobnie w tym samym czasie co pierwsza roślin lądowych. W skamieliny z florą Rhynie (np aglaofiton (w) , Rhynia (w) ), w wieku około 400 milionów lat zawierać morfologicznie identyczne mikoryza Glomales , związane z kłącza . Sugeruje to, że mikoryzy były instrumentem przyspieszonej kolonizacji wynurzonych ziem, dzięki ich zdolności do wydobywania wody i minerałów z gleby.
Te pierwsze skojarzenia pozwoliły również na ukonstytuowanie się gleby lepiej odpornej na złą pogodę, lepsze magazynowanie wody, przy jednoczesnej poprawie odporności roślin na stres wodny i zimno lub brak azotu (np. w świerku), ich odporność na wiatr i odpływu, a być może ich odporności na nadmierne natężenie światła, jak to ma miejsce w przypadku mykofikobiontów niektórych glonów przybrzeżnych (symbiozy te, głównie z workowcami , wydają się pojawiać wtórnie).
Asocjacje mikoryzowe (arbuskularne i ektomikoryzowe) występują również w zimnych ekosystemach (średnia temperatura poniżej 15°C), gdzie są aktywne przez cały rok lub część roku (w zależności od temperatury gleby), uczestniczą w pewnych adaptacjach odporności na żelowanie i promują przyswajanie składników odżywczych w niskiej temperaturze w grzybach mikoryzowych. Niewątpliwie przyczyniają się one do lepszego przeżywania grzyba w glebach zamarzniętych zimą, gdzie mikrogrzyby podlegają temu dodatkowemu czynnikowi selekcyjnemu. Z tych zimnych symbioz korzystają również trawy, takie jak jęczmień ( Hordeum ), który w ten sposób poprawia jego dostęp do fosforu w glebie .
Te symbiozy odpowiadają „ skokowi makroewolucyjnemu ” (skokowi ewolucyjnemu), ponieważ funkcje jednego partnera są powiązane z funkcjami drugiego, z mnożącymi się efektami funkcjonalnymi. W rzeczywistości przechodzimy ze stanu glonów i grzybów do stanu ziemskiej rośliny mikoryzowej (co jest sprzeczne z darwinowskim gradualizmem ).
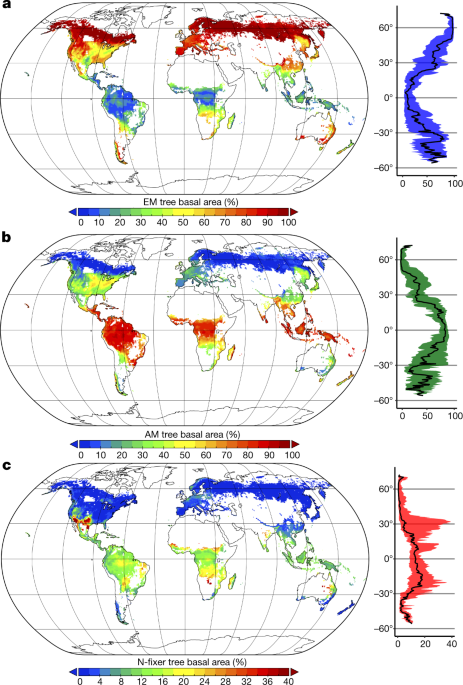
Obecnie 85% Archegoniatów , podobnie jak Hepatic , jest endomikoryzowych przez Glomales . Zakłada to, że symbioza z Glomalami jest najstarsza wśród Archegoniatów i że pozwoliłaby na imponujące promieniowanie tych ostatnich (różnorodność, lignifikacja …). 80% roślin naczyniowych jest skolonizowanych przez grzyby endomikoryzowe, podczas gdy ektomikoryzy są często związane z drzewami drzewiastymi.
Inne rodziny Glomeromycetes ( Acaulosporaceae i Gigaspora ) pojawiły się później około -250/-230 milionów lat temu. Mają doskonałe zdolności do eksploatacji zasobów mineralnych gleby.
Tymczasem ektomikoryzy mogły pojawić się w kredzie , ale najstarsze znane skamieniałości pochodzą dopiero z eocenu . Umożliwiły kolonizację wcześniej niesprzyjających gleb. Wtedy też pojawiły się symbiozy wiążące azot . Symbioza ektomikoryzowa występowała wielokrotnie niezależnie w różnych kladach grzybów (80 razy w Glomeromycetes , Ascomycetes i Basidiomycetes ) i Spermaphytes (12 razy w nagonasiennych i okrytonasiennych ). Ta częstotliwość pokazuje sukces ewolucyjny tych związków od przodków grzybów ektomikoryzowych, które żyły z martwej materii organicznej w glebie w saprofityczny sposób życia . Badania porównawcze genomów obecnych grzybów ektomikoryzowych i saprofitycznych pokazują, że komórki mikoryzowe utraciły wiele egzoenzymów, które zapewniają autonomiczne, węglowe odżywianie saprofitów, ponieważ są one odżywiane cukrami przez roślinę żywicielską, ale zachowały enzymy (peroksydazy, lakazy, fenolooksydazy grzybowe (np. tyrozynazy) wykorzystujące zasoby azotu i fosforanów pozostałości organicznych (lignina, związki fenolowe, celuloza) niezbędnych do prawidłowego funkcjonowania rośliny.
Niektóre rośliny z nowszymi symbiozami mikoryzowymi (z ewolucyjnego punktu widzenia) mogą również mieć związek z mikoryzami arbuskularnymi. Wydaje się więc, że w tego typu symbiozie pojawiły się ewolucyjne innowacje. Innowacje te niewątpliwie wyjaśniają ograniczoną liczbę roślin zdolnych do ich kurczenia się.
Pojawienie się ektomikoryz zostało dwukrotnie skorelowane z promieniowaniem ewolucyjnym roślin:
Ostatnio Pojawiły się jeszcze inne formy endomikoryzy i ektomikoryzy plutonu, zwłaszcza u Ericales , ze zjawiskiem mikoheterotrofii : ich symbionty grzybowe mają jeszcze silniejsze zdolności saprofityczne, co pozwala roślinie na ponowne wchłanianie węgla organicznego bezpośrednio przez grzyby, w gleby, w których aktywność mineralizacji jest bardzo niska ( wrzosowiska , wysokie góry, chłodne ekosystemy itp.): symbioza umożliwia łączenie poziomów troficznych .
Kilka grup roślin (mniej niż 10%) nie jest mikoryzowych. Wtórnie utraciły skojarzenie i ilustrują zjawisko neotenii . Są to zasadniczo rośliny pochodzące ze środowisk pionierskich (brak grzybów) lub bogate. Na przykład pianki przystosowane do nieprzyjaznych środowisk, w których mogą wysychać i gdzie grzyb może źle przeżyć. Posiadają anatomiczne struktury, które pozwalają im oprzeć się wysuszeniu, ale odpowiednio ich rozmiar pozostaje niewielki. Są to również rośliny okrytonasienne ( Brassicaceae , Chenopodiaceae , Polygonaceae , Proteaceae , Caricaceae itp.), które najczęściej przystosowały się do bogatych środowisk, w których zaopatrzenie hydromineralne nie wymaga mikoryz.
Mikoryza są pochodzenia najbardziej złożonych ekosystemów , a zwłaszcza w lasach , w szczególności lasów tropikalnych , które często żyją i ewoluować na glebach niewdzięcznych i czasami niepłodnych. Ich grzybnie tworzą wzajemnie połączone sieci grzybni, które wpływają na funkcjonowanie ekosystemów (cykle biogeochemiczne, skład zbiorowisk roślinnych, węglowe odżywianie siewek podczas ich rozwoju, modyfikacja konkurencji itp.) poprzez umożliwienie lub zwiększenie znacznych przepływów węgla organicznego i minerałów ( azot , fosfor , woda…) przez glebę (średnio 30 do 40% minerałów wychwyconych przez brzegi sieci grzybni jest zawracanych do korzenia, przy czym ten ostatni dostarcza 20 do 40% węglowodanów fotosyntezowanych do grzyba). Stanowią jeden z najbardziej dynamicznych elementów symbiozy mikoryzowej i odgrywają zasadniczą rolę w funkcjonowaniu i budowie zbiorowisk roślinnych. Transfery te są tak skuteczne, że podważają koncepcję specjacji poprzez rywalizację o składniki pokarmowe między roślinami ekosystemu, w szczególności o wychwytywanie fosforanów przez korzenie (pozwalają zrezygnować z nawozów fosforowych), o odporność na toksyczny wapń (niektóre rośliny wapniowe tolerują wapń dzięki strzępce grzybni aktywnie odrzucającej jony Ca 2+ na zewnątrz lub poprzez ich immobilizację w postaci kryształów szczawianu wapnia, które wytrącają się w wakuolach , idioblastach lub na ściankach strzępek) lub odporności na suszę . Są one jednak nadal mało wykorzystywane w ogrodnictwie , rolnictwie i leśnictwie lub do odkażania niektórych zanieczyszczonych gleb.
Pewne grupy grzybów są prawdopodobnie gatunkami kluczowymi, a nawet „gatunkami inżynieryjnymi”, które wpływają na główne procesy ekologiczne w glebie. Uważane są przez gleboznawców za istotny element różnorodności zbiorowisk, która jest czynnikiem stabilności i równowagi ekologicznej. Wiele kluczowych grup występujących w glebie (w szczególności bakterie i grzyby mikoryzowe) może łączyć się z roślinami (co najmniej 90% rodzin roślin lądowych) poprzez związki mikoryz arbuskularnych i odgrywać synergie niezbędne dla przetrwania i produktywności roślin, pomagając w tworzeniu zasadniczo pod ziemią sieć ekologiczna , której niektórzy biolodzy nazywają internetowej drewno szerokości (w odniesieniu do „ World wide Web ”). W 1997 roku kanadyjska badaczka Suzanne Simard jako pierwsza zademonstrowała tę sieć z mikoryzowym transferem węgla między drzewami w naturalnych warunkach.
Podejrzewa się, że większość grzybów mikoryzowych ma kilku żywicieli lub nawet szeroką gamę żywicieli (każda roślina kojarzy się powszechnie z kilkudziesięcioma różnymi grzybami mikoryzowymi), co wydaje się potwierdzać w środowiskach naturalnych , ale przeprowadzone na nich badania pokazują jednak, że uprawiane gleby uprawne , że różnorodność grzybów mikoryzowych jest „ekstremalnie niska w porównaniu z glebami leśnymi” .
Kolonizacja systemów korzeniowych, potencjał „mikoryzogenny” gleby i „zależność mikoryzowa” roślin są odwrotnie skorelowane z zawartością jonów fosforanowych w roztworze glebowym ; co więcej, wynik ten nie jest związany z żadną formą nawozu fosforowego, czy to organicznego czy nieorganicznego, ponieważ rośliny absorbują tylko jony w roztworze. Wzbogacenie tego roztworu staje się bezpośrednio odpowiedzialne za to, że dobrze odżywiona roślina nie sprzyja już rozwojowi mikoryz. W pewnych sytuacjach osiągnięty poziom fosforu staje się niekompatybilny z instalacją mikoryz.
Mikoryzy wchodzą również w interakcje z różnymi bakteriami glebowymi (m.in. Pseudomonas ), które mogą być chorobotwórcze (mikoryza chroni roślinę przed jej patogenami, np. emitując antybiotyki), ale które są również nazywane „bakteriami pomocniczymi do mikoryzacji” (w języku angielskim MHB: Mycorrisal Helper Bacteria ), ponieważ odgrywają one tak ważną rolę.
Mikoryzy wchodzą również w interakcje z innymi mikoryzami i innymi grzybami oraz z niektórymi powietrznymi drapieżnikami i pasożytami roślin:
Liczne kontrolowane eksperymenty z mikoryzacją wykazały, że naturalna lub sztuczna regeneracja korzysta z obecności lub inokulacji grzybami symbiontowymi dostosowanymi do roślin i kontekstu. Odwrotnie, kultura trufli mogła być eksportowana do Nowej Zelandii tylko z roślinami mikoryzowymi. Podobnie europejskie choinki rosły tam tylko ze swoimi symbiontami. Niektóre symbionty są spektakularnie skuteczne: jest to 60-procentowy wzrost całkowitej miąższości Pseudotsuga menziesii, który w ciągu 10 lat dopuszczono do zalesiania z roślin ektomikoryzowych w szkółkach szczepem Laccaria bicolor znanym jako S238N, w porównaniu z drzewami niezaszczepionymi.
Środki ostrożności : Istnieje ryzyko konkurencji z lokalnymi gatunkami, a nawet zanieczyszczenia genetycznego . Kilka obserwacji wykazało, że rośliny zaszczepione w szkółkach generalnie straciły tego symbionta na rzecz innych z lokalnej gleby, ale nie zawsze tak jest.
Wbrew powszechnemu przekonaniu związek mikoryzowy nie jest wyłącznie symbiotyczny, ponieważ transfery substancji z grzyba do rośliny nie zawsze zwiększają wartość selektywną (która jest kryterium definiowania symbiozy ). Korzyści, które różnią się znacznie w zależności od genotypu partnerów i środowiska, są trudne do oszacowania, zwłaszcza w przypadku roślin wieloletnich. Mogą korzystać z symbiozy tylko przez krótkie okresy w różnych okresach swojego życia. W rzeczywistości istnieje kontinuum relacji między partnerami, od symbiozy do pasożytnictwa poprzez saprotrofię (charakter relacji można opisać jako funkcję stosunku kosztów do korzyści).
Na przykład Tricholoma matsutake jest symbiotyczny, pasożytniczy lub saprofityczny w zależności od etapu rozwoju i warunków środowiskowych; storczyk mikoryza pasożytują na roślinach drzewiastych i gniazdo neotti ptasich pasożytują na ich mikoryz.
W rzeczywistości mikoryzacja jest stale niestabilna z powodu konfliktu interesów między partnerami i presji selekcyjnej, która popycha każdego partnera do bycia jak najbardziej pasożytniczym, co wyjaśnia wiele możliwych interakcji: wymianę składników odżywczych (roślina dostarcza cukrów i lipidów , grzybnia grzybów dostarcza wodę i sole mineralne, które czerpią z gruntu, czasem nawet kilkanaście metrów od korzenia, czynniki wzrostu ( hormony , witaminy ), allomony zapewniające ochronę. Nadmiar pasożytnictwa może doprowadzić do rozpadu związku. Z czasem pojawiła się „symbioza” i kilka razy zostałaby zerwana: skojarzenie jest odwracalne. Grzyby saprofityczne mogą zatem być symbiontami, które utraciły swojego żywiciela. Tak więc w niektórych skojarzeniach mikoryzowych jeden z partnerów wydaje się wykorzystywać drugiego (z natury lub w zależności od warunków). Pod pojęciem „wyzyskiwacz” rozumie się tutaj osobę, która uzyskuje korzyść (wzrost wartości selektywnej) bez wzajemności. Jako przykład możemy przytoczyć rośliny mikoheterotroficzne (ponad 200 gatunków znanych z storczykowatych , gentianaceae , burmanniaceae itp.), rośliny o zerowej lub zmniejszonej zdolności fotosyntezy, które pozyskują węgiel od swojego mikoryzowego partnera, który sam pozyskuje go od „partnera chlorofilowego”. przez sieć grzybni.
Wyzysk jednego z partnerów symbiozy przez drugiego zakłada rekompensatę dla wyzyskiwanej jednostki:
Mechanizmy kompensacyjne mogą być ważne w stabilizacji symbiozy. Ogólnie rzecz biorąc, symbiozy charakteryzują się spadkiem dryfu genetycznego i szybkości specjacji (por. efekt „Red King” ), ale nie zawsze tak jest w przypadku mikoryzacji: im bardziej związek jest eksploatacyjny, tym bardziej jest specyficzny, ponieważ wyzyskiwany rozwija opór, który wyzyskiwacz musi ominąć ( Teoria czerwonej królowej ). W niniejszym przypadku selektywna presja na gospodarza doprowadziłaby do pojawienia się ligniny i sprzyjała wzrostowi tkanek naczyniowych.
Wreszcie logika i wiele wskazówek sugeruje pasożytnicze pochodzenie mikoryz:
Istnieją dwa główne typy mikoryz, definiowane przez relacje fizjoanatomiczne między obydwoma partnerami: endomikoryza i ektomikoryza. Ericoid mikoryza i te związane z orchidei są również badane na ich usług ekologicznych , ale mają bardziej ograniczony interes ekonomiczny.
Globalne mapowanie symbioz leśnych pokazuje ewolucyjny sukces drzew ektomikoryzowych. Z trzech najbardziej popularnych typów symbiozie (arbuscular endomycorrhizae bogate w gorącym i wilgotnym klimacie tropikalnym, ectomycorrhizae w zimnych klimatach i azot- mocujących bakterii w suchych i gorących klimatach) stanowią ectomycorrhizae tylko 2% gatunków roślin, a 60% stanowią wszystkie drzewa na planecie.
Najczęstszą postacią są endomikoryzy (lub mikoryza wewnętrzna). Są to mikoryzy, które wnikają do wnętrza korzeni, aby lepiej się z nimi kojarzyć.
Istnieje kilka rodzajów endomikoryz:
Kojarzone są z roślinami zielnymi i drzewiastymi. Te endomikoryzy arbuskularne, zwane także mikoryzami pęcherzykowymi i arbuskularnymi (lub mikoryzami pęcherzykowo-arbuskularnymi), biorą swoją nazwę od pęcherzyków międzykomórkowych (często krople oleju, rezerwa zapasowa dla grzyba) i struktur „wewnątrzkomórkowych” przypominających małe drzewo. Jeśli jednak dobrze przechodzą przez ścianę, nie penetrują błony plazmatycznej komórki roślinnej, co może powodować inwazję jej błony. Ma to wpływ na zwiększenie powierzchni kontaktu między strzępką a komórką roślinną, a tym samym ułatwia wymianę metabolitów między dwoma partnerami.
Endomikoryzy arbuskularne są tworzone tylko przez grzyby z grupy Glomeromycetes, które utraciły zdolność rozmnażania płciowego. Strzępki rozciągają się do miąższu korowego korzenia, tworząc pęcherzyki zawierające rezerwy i struktury rozgałęzione, arbuskuły. Dlatego rozmnażają się tylko bezpłciowo. Jednak strzępki różnych osobników mogą się łączyć, umożliwiając wymianę genetyczną i formę paraseksualności.
Są również unikalne genetycznie, ponieważ ich zarodniki mają kilka genetycznie różnych jąder.
W tej symbiozie gęsta i rozległa sieć strzępek grzybów mikoryzowych pomaga roślinie eksplorować zwiększoną objętość gleby i uzyskiwać dostęp do miejsc niedostępnych dla korzeni. Grzyb pozwala roślinie poprawić odżywianie poprzez dostarczanie głównie wody, fosforu i azotu . Nawiasem mówiąc, kolonizacja korzeni przez grzyby mikoryzowe umożliwia ich ochronę przed atakami organizmów chorobotwórczych. Kilka badań wykazało, że bez skojarzenia z grzybem symbiotycznym roślina będzie rosła wolniej (jeśli w ogóle, jak w przypadku sosny czarnej na glebie wapiennej) i będzie znacznie bardziej narażona na infekcję. W zamian grzyb korzysta z fotosyntezy rośliny w postaci bogatej w energię materii organicznej ( cukry ) niezbędnej do jej przetrwania. W ten sposób poprawia się jakość próchnicy , z korzyścią dla innych gatunków oraz dla utrzymania lub budowy i poprawy gleby.
Wiadomo, że niektóre rośliny z rodziny Gymnosperm , takie jak Podocarpaceae , Araucariaceae i Phyllocladaceae , mają obrzęki korzeni atakowane przez grzyby endomikoryzowe, zwane myconodules lub pseudogules .
W ectomycorrhizas (lub mikoryza zewnętrzny) dotyczą jedynie 5% roślin naczyniowych, głównie drzewa strefy umiarkowanej i lasów borealnym (takich jak Fagaceae , w Pinacées lub Betulaceae ) oraz grzyby z podziałem Ascomycetes , w Basidiomycetes i Zygomycetes . Te mikoryzy nie przenikają przez ściany komórkowe wewnątrz komórek rośliny, ale po prostu otaczają korzenie, tworząc płaszcz grzybni i sieć między ścianami komórkowymi korzenia.
Grzyb najpierw kojarzy się z drobnymi korzeniami o zdecydowanym wzroście, pozbawionymi włośników. Następnie otacza korzeń płaszcza strzępek , rękawa mikoryzowego. Inne strzępki rosną między komórkami w zewnętrznej części miąższu korowego, tworząc symbiotyczny interfejs lub „ sieć Hartiga ”. Symbioza modyfikuje fizjonomię korzenia mikoryzowego: pęcznieje, przestaje rosnąć i może obficie się rozgałęziać. Czapeczka i merystem wierzchołkowy są następnie redukowane.
Setki różnych grzybów mogą być powiązane z tym samym gatunkiem. Na przykład buk ma „rekord” z ponad 200 skojarzeniami mychoriziennes.
Wzrost grzybni w kierunku korzeni rośliny żywicielskiej jest wywoływany przez percepcję sygnału emitowanego przez korzenie w postaci cząsteczek specyficznych dla rośliny, takich jak strigolaktony , a zwłaszcza flawonoidy .
Sygnały emitowane przez grzyba, które pozwalają na jego rozpoznanie przez roślinę, są słabo poznane. Przez analogię do czynników Nod, czynniki te nazywane są „czynnikami Myc”. Ostatnio wykazano, że cząsteczki czynnika Nod wytwarzane przez grzyby arbuskularne i mające wpływ na roślinę żywicielską, ich rola w sygnalizacji symbiotycznej pozostaje do ustalenia.
Szlak przekazywania sygnałów myc czynnik aktywny zawiera etapy wspólnego z obecnej ścieżki Nod z brodawek roślin motylkowych o Rhizobium i w actinorrhizal roślin . Receptory czynników Myc nie są znane, ale białka interweniujące później, takie jak SYMRK, CASTOR, POLLUX, NUP, CYCLOPS, są niezbędne do symbiozy ryzobialnej i arbuskularnej mikoryzy. Podobnie jak w przypadku symbiozy ryzobialnej, kontakt z grzybem arbuskularnym indukuje oscylacje wapnia. Ponieważ endomikoryza arbuskularna pojawiła się przed endosymbiozą wiążącą azot, naukowcy stawiają hipotezę, że szlak transdukcji sygnału mikoryzowego był rekrutowany przez bakterie wiążące azot.
Optymalna procesja mikoryzowa działałaby jak „nawóz” bez wad i bezpłatnie, bez pestycydów , fungicydów i poprawek . Po wstępnych próbach w Australii w latach 20. XX wieku firmy opracowały szczepy do zaszczepiania odpowiednie dla kilku gatunków handlowych (pojedyncze lub wieloszczepowe inokula), ale musiałyby również zostać dostosowane do kontekstu lokalizacji. Kilku badaczy uważa, że szczepy genetycznie symbiotycznych grzybów, które najlepiej nadają się do pracy w terenie, to te, które są rodzime. Prywatne firmy na całym świecie rozwinęły w ten sposób produkcję i sprzedaż modyfikatorów mikoryzowych przeznaczonych do użytku rolniczego ( bionawóz , biostymulator ), ale także ekologicznego ( odnowa gleb zerodowanych, fitoremediacja ).
W rolnictwie zastosowanie grzybów mikoryzowych pozwoliłoby uniknąć wielu problemów związanych z denaturacją gleb organicznych. W rzeczywistości dodanie pestycydów, aw szczególności fungicydów, może mieć szkodliwy wpływ na jakość gleby. Grzyby odgrywają ważną rolę w utrzymaniu żyznej gleby. Dodatek związków chemicznych o wysokiej zawartości fosforu spowodowałby zmniejszenie ilości i jakości grzybów endomikoryzowych, zmniejszając dostarczanie składników odżywczych do roślin uprawnych. Ponadto zbyt duża ilość fosforu ma negatywny wpływ na drogi wodne (eutrofizacja). Dlatego ważne jest, aby zmniejszyć ilość szkodliwych chemikaliów i zamiast tego zdecydować się na szczepienie naturalnymi grzybami, które miałyby te same zalety szybkiego wzrostu, bez negatywnych skutków dla środowiska.
Badanie na drzewie jujuby wykazało, że kontrolowana mikoryzacja była korzystna dla jego wzrostu i odżywiania fosforanami. Rzeczywiście, drzewo owocowe znacznie łatwiej przyswajało naturalny fosfor dodawany na ubogiej glebie. Był nawet zależny od tego dla normalnego wzrostu. Dlatego w rolnictwie nie wystarczy po prostu dodać fosforu w bardzo dużych ilościach, aby roślina rosła szybciej, to także kwestia uwzględnienia sieci mikoryzowej. Musi przede wszystkim być w stanie skutecznie się wchłaniać. Przedawkowanie składników pokarmowych niczego by nie zmieniło, a to spowodowałoby jedynie wypłukiwanie fosforu i zanieczyszczenie wód powierzchniowych.
Działanie mikoryz nie kończy się na lepszym odżywieniu: pozwalają one również roślinie lepiej chronić się przed szkodliwymi czynnikami biotycznymi i abiotycznymi. Oczywiście lepsze odżywianie oznacza lepsze zdrowie roślin, które mogą wtedy lepiej tolerować stresy środowiskowe, w szczególności patogeny. Mikoryzy mogą uwalniać do ryzosfery antybiotyki, które eliminują wszystkie drobnoustroje chorobotwórcze lub nie, co może zmienić kierunek handlu na ich korzyść. Ponadto symbioza mikoryzowa powoduje również ustanowienie mechanizmów obronnych w samej roślinie (odporność i stymulacja immunologiczna). Jest to pośrednia ochrona, która znajduje się na poziomie komórkowym. W roślinach ektomikoryzowych komórki ryzodermy syntetyzują przechowywane w wakuolach garbniki, które zapewniają ochronę przed kolejnymi atakami drobnoustrojów i innych patogenów. Roślina produkuje również więcej ligniny w ścianach komórkowych endodermy i tkankach naczyniowych. Może również indukować lub tłumić różne mechanizmy obronne związane z fitoaleksynami , peroksydazami , chitynazami i kilkoma innymi. Krótko mówiąc, grzyb powoduje znaczne zmiany w mechanizmach obronnych rośliny przed różnymi pasożytami i to przed infekcjami.
Czasami skojarzenie z grzybem powoduje przebudowę systemu korzeniowego. Na przykład w truskawkach mikoryzacja powoduje zwiększoną ochronę przed zgnilizną korzeni powodowaną przez Phytophthora fragariae . Intensywnemu rozgałęzianiu się korzeni indukowanemu przez grzyby mikoryzowe towarzyszą wysięki z korzeni, co może skutkować modyfikacją mikroflory i bezpośrednimi interakcjami z pasożytami.
W przypadku grzybów pasożytniczych lepiej chroniona jest również roślina związana z mikoryzą. Strzępki symbiotycznego grzyba kolonizują korzenie rośliny i „blokują” miejsca dostępu dla pasożytniczego grzyba. Konkurują również z dostępnymi składnikami odżywczymi. Mikoryza również czerpie korzyści z wiązania się z rośliną: otrzymuje związki węgla w zamian za minerały. Ponieważ węgiel jest bardzo pożądany, mikoryza ma większą przewagę niż pasożytniczy grzyb wolny w glebie. Ponadto mikoryzy mogą stanowić do 80% masy mikrobiologicznej gleby. W związku z tym mają duży wpływ na fizyczne i chemiczne właściwości środowiska, dzięki czemu mogą kontrolować szereg interakcji drobnoustrojów w glebie. Czasami jest w zgodzie z mikoryzową rośliną zapewniając ochronę i wzrost, ale czasami efekty są raczej negatywne.
Mikoryza może zatem pomóc chronić roślinę przed stresami abiotycznymi (np. suszą) i biotycznymi na kilka sposobów: lepsze odżywianie i zdrowie, transformacja morfologiczna korzeni, indukcja lub tłumienie mechanizmów obronnych oraz poprzez oddziaływanie na samego pasożyta poprzez rywalizację o zasoby i miejsca infekcji lub przez modyfikację mikroflory i wzrost poziomu materii organicznej.
Należy jednak zauważyć, że te interakcje są badane w szklarniach iw kontrolowanych środowiskach i dlatego nie odzwierciedlają w pełni całej złożoności środowiska naturalnego, które zmienia się w czasie i w składzie.
Biorąc pod uwagę wszystkie wymienione zalety mikoryz, można śmiało powiedzieć, że ich właściwe stosowanie pozwoliłoby uniknąć wielu problemów środowiskowych, ponieważ działałyby jako nawozy i środki ochronne. Stosowanie nawozów chemicznych, fungicydów i pestycydów nie byłoby wtedy konieczne. Rzeczywiście, nawozy zmniejszają tempo mikoryzacji, co skutkuje zwiększoną zależnością roślin od tych składników. Podobnie stosowanie pestycydów zmniejsza siłę zaszczepiania gleby, co skutkuje zmniejszeniem ochronnego działania mikoryzy i zwiększonym zapotrzebowaniem roślin na pestycydy. Dodanie takich danych wejściowych tworzy formę błędnego koła .
Działalność leśna i rolnicza może zaburzać lub modyfikować (negatywnie lub pozytywnie) mikroflorę grzyba i jego zdolność do mikoryzacji;
Zmiany biochemiczne i biologiczne, a także zmiany mikroklimatyczne wywołane dużymi zrębami zupełnymi (lub nawet zbiorami w przypadku rolnictwa) mogą mieć niedoceniany wpływ, związany z bezpośrednim oddziaływaniem na glebę, wodę, ale także z utratą grzybów. inokulum (nawet jeśli korzenie pozostają w glebie, „ duże drewno ” i duże „ martwe drewno ” stają się rzadkie lub nieobecne i często zbiera się cięcie).
Termin mikoryza (od greckiego miko , „grzyb” i rhiza „korzeń”) został wprowadzony w 1885 roku przez botanika Alberta Bernharda Franka . Wynajęty przez króla pruskiego Wilhelma I st. do opracowania metod produkcji trufli poprzez badanie ich rozprzestrzeniania się, ABFrank zauważa, że grzyby te kojarzą się z korzeniami drzew i proponuje, że jest to związek symbiotyczny, a nie pasożytniczy . Teoria Franka o ekologii ewolucyjnej , zaprzeczająca przekonaniu, że trufle i inne grzyby powodują choroby roślin i gnicie, jest gorąco kwestionowana przez jego kolegów. Teoria ta powraca na centralne miejsce wraz z pracami Jacka Harleya uważanego za „papieża mikoryzy” dla swoich studiów od lat pięćdziesiątych, Barbary Mosse w 1962 roku, JM Phillipsa i DS Haymana w 1970 roku, M. Gardesa i TD Brunsa w 1993 roku.
Badania szybko rozwijające się na początku XXI -go wieku z nowych środków biologii molekularnej i genetyki do punktu podając nazwę do dyscypliny naukowej , w mycorhizologie , studiował przez mycorhizologues . W latach 2008-2016 ponad 10 000 nowych prac naukowych dotyczyło mikoryz.
{kind=link}